|
|
一開始,分子生物學家做出來的結論比較支持大麥起源於西亞的傳統說法。比如2000年一項由埃及和德國學者共同完成的研究就顯示,大麥很可能起源於今以色列和約旦之間的一個狹窄的地域[6]。然而,隨著科學家使用的核酸片段不斷增長,結論也發生了變化。還是在2005年,就有學者通過比較大麥的葉綠體DNA發現,現在的大麥可能是多次起源的,也就是說,它在不同的地方被馴化了不止一次。除了西亞之外,青藏高原也可能是一個獨立的起源地[7]。
要想解決這個問題,只能是採集更多的野大麥和栽培大麥的標本、比較更長的核酸片段。以色列海法大學長期從事演化生物學研究的學者內沃(Eviatar Nevo)便和浙江大學的張國平合作,內沃提供了西亞的大麥標本,而張國平則找來了徐廷文等人花費幾十年功夫從青藏高原各地採集的大麥標本。他們在全基因組的範圍內比較了大麥的DNA「指紋」,在2012年初步認定大麥是多次起源的。首先,他們發現雖然青藏高原的野大麥有豐富的遺傳多樣性,但終歸比不過西亞的野大麥居群——畢竟大麥們是從約276萬年前開始從西亞一步步向東,到達青藏高原北沿之後才慢慢「爬」上世界屋脊的。其次,他們發現西亞的栽培大麥的DNA「指紋」非常接近西亞的野大麥,而青藏高原和中原地區的栽培大麥的DNA「指紋」卻非常接近青藏高原的野大麥,這似乎意味著西亞人的先民在以色列、約旦一帶,藏族的先民在青藏高原先後獨立地馴化了大麥,從而形成了栽培大麥的西、東兩大支系[8]。

青藏高原上的青稞田。圖片:shutterstock.com
然而,正如栽培水稻會和野生稻發生基因交換,造成水稻多次起源的假象一樣,無論是西亞還是青藏高原,栽培大麥的基因也都可能和野大麥的基因發生「漸滲」,造成大麥多次起源的表面現象。2008年,日本學者就發現大麥的帶皮和裸粒性狀差異僅僅由一個基因控制,而裸粒的基因形式是單起源的。這樣來看,大麥是單次起源還是多次起源,仍然是一個懸而未決的問題。
不過,如果結合考古證據來看的話,大麥西來的可能性要更大一些。1977年在西藏昌都發現了卡若遺址,定年為相當於中原龍山時代的5000–4000年前。卡若遺址中出土了大量穀物遺存,經鑒定卻全都是粟(小米),不雜一點其他穀物,說明在這個時候,藏族先民還沒有種青稞。然而在1994年,在雅魯藏布江河谷附近發現了定年為約3500年前的昌果溝遺址,其中卻不僅有粟,有青稞,還有零星的小麥遺存。考古人員懷疑小麥只是混雜在青稞中的雜谷,既然小麥是西來的穀物,那麼夾雜著小麥的青稞很可能也是西來的穀物[9]。事實上,如果對整個青藏高原的新石器時代遺址的植物遺存情況進行匯總,就會發現青稞基本上是約3600年前才突然出現的,並沒有經歷西亞那樣漫長的馴化過程,這也強烈暗示了青稞從西方傳來[10]。
儘管如此,經過和青藏高原野大麥的長期雜交,青稞早就吸收了大量本地野大麥的優良基因,從而適應了世界屋脊的嚴酷環境。更有趣的是,內沃和張國平用大麥的轉錄RNA進行的另一項研究還發現,如果除去大麥基因組中的大量「垃圾」片段,僅就能夠翻譯成蛋白質、直接決定大麥的各種具體性狀的基因而言,西亞的野大麥和青藏高原的野大麥對全世界栽培大麥(青藏高原上的青稞除外)的基因的貢獻竟然差不多大。非要說區別的話,在大麥的7對染色體中,西亞野大麥對其中編號為1–3的3對染色體基因的貢獻略多一些,青藏高原野大麥對編號為4–7的4對染色體基因的貢獻略多一些[11]。也就是說,大麥不僅曾沿著史前的青銅之路和遠古的絲綢之路從西向東傳播,而且還曾頻繁地從東向西傳播,這樣才讓現代栽培大麥的基因既有西亞野大麥的貢獻,又有青藏高原野大麥的貢獻。一言以蔽之,栽培大麥是單次起源還是多次起源並不重要,重要的是它很早就融合了東西雙方的優良基因。
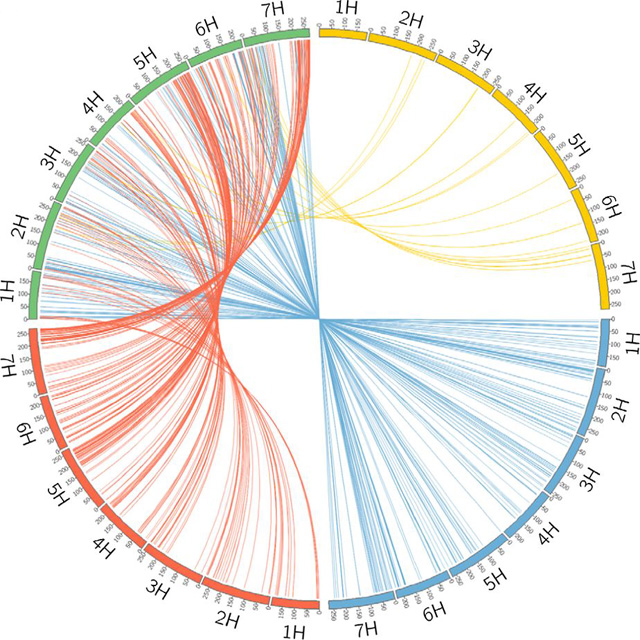
栽培大麥(綠色)、青藏高原野生大麥(紅色)與兩種近東地區大麥(藍色、黃色)的基因組比對。圖片:參考文獻11
這樣的結果真可謂充滿了隱喻性。要知道,藏族這個高原民族本身也是混合起源。藏族民間一直廣泛流傳著一個有關藏族起源的故事:說一隻孜孜修行的公獼猴最終同意了化妝成母猴的岩羅剎女(「羅剎」是惡魔的意思)的求愛,在觀世音菩薩的准許下與她結為夫妻,並生下了六隻小猴。誰知被送進樹林中生活的小猴很快就繁衍出很多後代,把樹林里的果子都摘光了,不得不餓肚子,老猴於是把它們領到一處長滿野生穀物的山坡,讓它們以野谷為食,眾猴吃了野谷之後,身上的毛便慢慢變短,尾巴也漸漸消失,漸漸學會了說話,成了人類的祖先。對於這則傳說,有藏族學者就從人類學的角度考證,認為反映了藏族由山地部族(獼猴)和森林部族(岩羅剎女)混合形成的族源[12]。

拉薩,一位藏族婦女正在收穫青稞。圖片:123rf.com
分子人類學研究確認,藏族的確是典型的混合起源民族。話說晚期智人在約7萬年前走出非洲之後,其中一支可能沿著亞洲大陸南部的海岸線一路到達今印度東部。大概就是在這個地區附近,晚期智人遇到了上一次走出非洲的早期智人海德堡人的後代——丹尼索瓦人,並和他們發生了混血。通過這次混血,這一支晚期智人幸運地獲得了能夠適應青藏高原稀薄氧氣的基因形式,這使他們得以在1.5–2萬年前攀上高聳的青藏高原,靠著這天賦異稟在其他人群難於生存的地方生活[13]。
後來,晚期智人的另一支也在約3萬年前到達東亞,並很快佔據了東亞大陸的大部分地區,成為今天漢族人父系的主要來源。然而在大約5200年前,其中的一個小支從黃土高原西麓也攀上了青藏高原。這批後來的居民具有高超的農業技術,先是把中原的粟和黍、繼而又把西來的小麥和青稞帶到高原之上。通過種植和野大麥雜交后更為耐寒的青稞,他們一步步向高原深處挺進,與高原上仍然進行狩獵–採集的原住民混合,最終便形成了藏族的先民。今天的藏語和漢語之所以有一定親緣關係,在語言學上同屬於漢藏語系,也正是因為這進入高原的后一支系在擴散的過程中保住了自己的語言的基本框架。可以說,沒有先後兩次的族群融合和由此造成的文化融合,藏族人就不可能把世界屋脊當做自己的家園,成為其他民族只能望其項背的高原雄鷹。
其實何止是藏族,在分子人類學家眼中,漢族本身又何嘗不是古代的許多族群混合之後的產物呢。在這些族群接觸的時候,當然不可避免會有殘酷的競爭和殺戮,但無論如何,我們終究是這些混合族群的後裔,而且在今天理所當然地分享著由這些不同的族群文化彼此「涵化」之後形成的混合文化。
不管你在民族觀念上是什麼樣的態度,這些都是現代生物學揭示的無法迴避的事實。(編輯:老貓)參考資料Sencer HA et Hawkes JG (1980) On the origin of cultivated rye. Biol. J. Linnean Soc. 13:299–313.鄭殿升, 張宗文 (2011) 大粒裸燕麥(莜麥)(Avena nuda L.)起源及分類問題的探討. 植物遺傳資源學報12:667–670.Blattner FR (2009) Progress in phylogenetic analysis and a new infrageneric classification of the barley genus Hordeum (Poaceae: Triticeae). Breed Sci. 59:471–480.Kislev ME, Nadel D et Carmi I (1992) Epipalaeolithic (19,000 BP) cereal and fruit diet at Ohalo II, Sea of Galilee, Israel. Rev. Palaeobot. Palynol. 73:161–166.中央民族學院少數民族語言文學系藏語文教研室藏族文學小組 (1980) 藏族民間故事選. 上海: 上海文藝出版社.Badr A et al (2000) On the origin and domestication history of barley (Hordeum vulgare).Mol. Biol. Evol. 17:499–510.Molina-Cano JL et al (2005) Chloroplast DNA microsatellite analysis supports a polyphyletic origin for barley. Theor. Appl. Genet. 110:613–619.Dai F et al (2012) Tibet is one of the centers of domestication of cultivated barley. Proc. Natl. Acad. Sci. U.S.A. 109:16969–16973.傅大雄, 阮仁武, 戴秀梅等 (2000) 西藏昌果古青稞、古小麥、古粟的研究. 作物學報 26:392–398.Chen FH et al (2015) Agriculture facilitated permanent human occupation of the Tibetan Plateau after 3600 B.P. Science 347:248–250.Dai F et al (2014) Transcriptome profiling reveals mosaic genomic origins of modern cultivated barley. Proc. Natl. Acad. Sci. U.S.A. 111:13403–13408.袁建勛, 才貝, 卓遜·道爾吉 (2005) 藏族起源的「金枝」——解讀「獼猴與羅剎女結合」. 西北民族大學學報(哲學社會科學版) 2005(4):116–119,128.Huerta-Sánchez E et al (2014) Altitude adaption in Tibetans caused by introgression of Denisovan-like DNA. Nature 512:194–197.
|
|





 狗仔卡
狗仔卡













 提升卡
提升卡 置頂卡
置頂卡 沉默卡
沉默卡 喧囂卡
喧囂卡 變色卡
變色卡 搶沙發
搶沙發 樓主
樓主
